研究の背景
呼吸は、これまで述べてきたセントラルコマンドまたは末梢神経反射による呼吸中枢刺激という反射的な調節
のほかに,骨格筋と同様に意識的・随意に変えることも可能です.最近,この経路が注目されてきました.
Finkらは,ポジトロン断層法(positron emission tomography; PET)を用い,脚運動時には,運動指令を発する
大脳皮質の一次運動野の脚運動に関係する部位とともに,同じ一次運動野の意識呼吸に関係する部位も活性化
しており,呼吸がすぐには低下しない運動後も,その意識呼吸に関係する部位の活性化が持続していることを
報告しています.つまり,運動時には,骨格筋と同じく,皮質-脊髄路(錐体路)を通って呼吸筋に指令を送るという,
意識的な換気調節が行われている可能性があります.
Yunokiらはこの点についてユニークな実験をしています.被検者に定常負荷運動と言いながら,負荷をゆっくり
ランプ状に上げていくと(騙し試行),負荷の変化の意識がほとんどなく,ランプ負荷と言ってランプ負荷運動させた
場合(通常試行)に比べ,換気が減少していたことから,中程度までの運動では,負荷の変化に対する
「意識」「注意」が換気に影響していることを示唆しています.
一方,セントラルコマンドや末梢神経反射がない運動前の安静状態でも、呼吸は予測的に増加し,運動をイメージ
したり,催眠術で運動を暗示しただけでも換気は増加することが報告されています.Williamsonらは,催眠術をかけて
運動をイメージさせると,努力感(effort sense)が高まるとともに,心拍数などの循環系が亢進することから,努力感が
セントラルコマンドと密接に関係し,運動時の循環応答に影響を及ぼすことを示唆しています.
さらに彼らは,セントラルコマンドには,上位中枢から活動筋への運動指令(セントラル・動作・コマンド)と上位中枢
から循環系への指令(セントラル・循環・コマンド)の二種類あり,それぞれ独立して働くという新しいセントラルコマンド説
を提唱しています.残念ながら彼らは呼吸について言及していませんが,運動時の循環応答も換気応答と同じ
セントラルコマンド説が関与しており,「循環」は「呼吸」に置き換えることが可能と考えられます.
すなわち,従来の呼吸中枢(および循環中枢)に放散するセントラル(動作)コマンド説の経路だけでなく,皮質-脊髄路を
通って直接的に呼吸筋に指令(セントラル・呼吸循環・コマンド)を送る経路でも,換気を亢進させていると考えられます.
これは上述したFinkらの結果と一致しています.
呼吸筋への出力経路だけでなく,出力を決定する入力と統合プロセスも重要です.随意運動,例えば野球のバッティング
の場合,ボールの速度や高さを「認知(知覚と判断)」し,これまでの「学習」「記憶」からタイミング,振る速さおよび高さを
決めて動作をプログラミングし,一次運動野から錐体路経由で各筋に命令を出力してボールを打っています.
同様に運動時の呼吸は,強度に合わせて随意的に呼吸筋を収縮させているとすると,まず運動強度を「認知」し,
これまでの「学習」「記憶」から呼吸の大きさを決定し,呼吸筋に運動指令を出力することが考えられます.
この運動強度の認知については,前述したようにイメージ運動では,努力感に合わせて呼吸・循環が高まることから,
強度そのものよりも運動の努力感(つらさ,きつさ)を認知していると考えられます.そして,以前の経験(記憶)から,
この「きつさ」ならこれだけ換気を上げたらよいということを学習しており,それに合わせて随意的に換気を上げている
のではないかと推察できます.
もしこの仮説が正しいならば,運動時の換気に「認知」や「学習・記憶」が関与しているはずです.運動時の換気応答に
認知機能が関与することを証明する実験として,Bellら は,パズルをしながら,すなわち認知(この場合はきつさの認知)を
妨げながら運動すると,換気増大が減弱することを報告しています.一方,学習・記憶が関与することを示す実験として,
Wuyamらは,トレッドミルの音を聞かせて運動をイメージさせると,一般人に比べよくトレーニングされた競技選手は換気が
増大することを報告しています.また,Thorntonらは,PETを用い,催眠術で急坂を自転車で登るイメージを想起させると,
換気の増大とともに前頭連合野,運動前野などの前頭葉や視床および小脳が活性化していることを報告しています.
前述したFinkらも運動中には意識呼吸の部位以外に前頭葉,視床,小脳,さらに大脳辺縁系や大脳基底核も活性化して
いたと報告しています.これらの部位は,運動制御とともに学習,記憶,認知,意識などと密接に関係しています.
以上のことから,運動時換気亢進には,前頭葉の運動関連領域や連合野,皮質下(大脳辺縁系,大脳基底核),
視床および小脳において,運動強度を努力感(つらさ)として認知し,学習・記憶から呼吸の大きさを決定し,
一次運動野の意識呼吸の部位から錐体路を経由して呼吸筋に出力する調節経路,すなわち「学習・認知による
意識的調節」が関与していることが強く推察されます.今のところ確証はありませんが,脳科学の進歩により,
これらの点について今後明らかにされていくことが期待されます.
実験成果
運動時の換気亢進については,上述したように、従来はセントラルコマンドと末梢神経反射の2つの神経性のメカニズムで
説明されてきましたが,2000年以降,「学習・記憶」が関与しているという新しい仮説(「学習仮説」)が提唱され,
生理学的常識を覆すものとして注目を浴びています.この学習効果が確かめられれば,効率的な呼吸法を「学習」させる
ことに繋がり,一般人の有酸素性運動に役立つと考えられます.
この仮説を検証するには,通常と異なる換気を学習させる必要があること,換気増大の時間相別に詳しく分析することが
必要ですが,これまでそのような研究は見当たりませんでした.
そこで我々は,2つの方法,すなわち胸郭制限と低酸素を用いて,運動時の換気を人為的に増やして短期間トレーニングさせ,
その後の通常の状態で運動させても,トレーニング前より換気が上がっているかどうかを確認し、学習仮説が正しいかどうか
確かめようとしました.
(1)胸郭制限による学習効果実験
方法
胸部をベルトで締めて胸郭制限下で運動すると,最高酸素摂取量(VO2peak)の60%以下の強度では,胸郭制限しない通常の
運動時に比べ,毎分換気量や努力感が増加することを,我々は先行研究(デサントスポーツ科学)で明らかにしました.
そこで,健康な大学生男子14名に対し,まずVO2peakの50%の負荷で6分間×5セットの自転車漕ぎ運動を,胸郭制限のない
通常状態で実施しました(Pre).毎分換気量,酸素摂取量(VO2),心拍数などについて,1秒値変換後に加算平均し,
運動開始20秒以内のPhase I,指数関数的増加部分のPhase II,定常状態のPhase IIIに分けて分析しました.
次に被検者7名については,ベルトで胸郭を締め,肺活量が30%程度低下する胸郭制限条件で,50%VO2peak の負荷で5分間の
運動を1日4セット,週5日2週間計40回実施した(CR群).トレーニング終了後(Post),胸郭制限なしでPreと同様の測定を
実施し,胸郭制限せずに同様にトレーニングした群(NR群)と比較しました.
結果 全体的傾向として,NR群ではトレーニングの初期効果(呼吸数の安定・低下)により,毎分換気量がトレーニング後に
低下するのに対し,CR群では逆にトレーニング後にやや増加する傾向が見られました(図1).
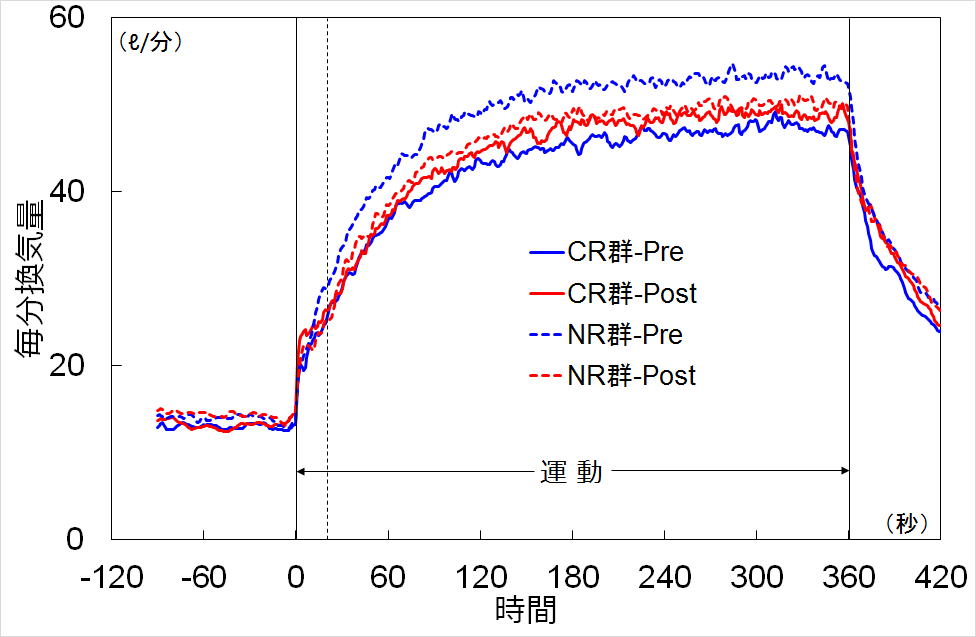
図1.胸郭制限トレーニング前後の毎分換気量の変化
時間相別に見ると,毎分換気量と呼吸数の運動開始20秒以内(Phase I)のゲイン(増加分)は,NR群ではPreと比べPostで
低下しますが,CR群では増加傾向を示し,毎分換気量のNR群とCR群で有意な交互作用が認められました.
また,毎分換気量のPhase IIの時定数,Phase IIIのゲインに両群で差は認められませんでした.
結論
このことから,運動開始直後(Phase I)の換気急増に学習効果が関与することが明らかとなり,それには努力感などの
上位中枢が影響していることが示唆されました.
(2)低酸素による学習効果
方法
低酸素を吸入しながら運動すると,常酸素吸入時に比べ毎分換気量は増加します.そこで,健康な大学生男女16名に対し,
まず,酸素濃度21%の常酸素条件下で50%VO2peakの負荷で5分間の自転車漕ぎ運動を十分な休憩をはさんで5セット実施し,
呼吸循環応答を(1)と同様に測定しました(Pre).
4週間以上経過した後,被検者8名については,常酸素で安静の後,運動開始時に吸入酸素濃度を13%に切り替え,
50%VO2peakの負荷で5分間の自転車漕ぎ運動を,7分の休憩をはさんで6セット繰り返すトレーニングを3日間連続で実施
しました(Hypo群).4日目にPre測定と同様に常酸素条件下での運動に対する呼吸循環応答を測定し(Post),同様の
トレーニングを常酸素で実施した群(Norm群)と比較しました.なお,被検者にはどの測定においても吸入ガス組成について
の情報を与えませんでした.
結果
図2に示すように,Hypo群ではトレーニング中(Tr)は低酸素吸入により,通常空気条件のトレーニング前(Pre)に比べ
換気が増大し,トレーニング後(Post)に常酸素で運動すると,同条件のPreに比べやや換気が増加していました.
一方,Norm群ではトレーニングにより毎分換気量は減少する方向に向かいました.
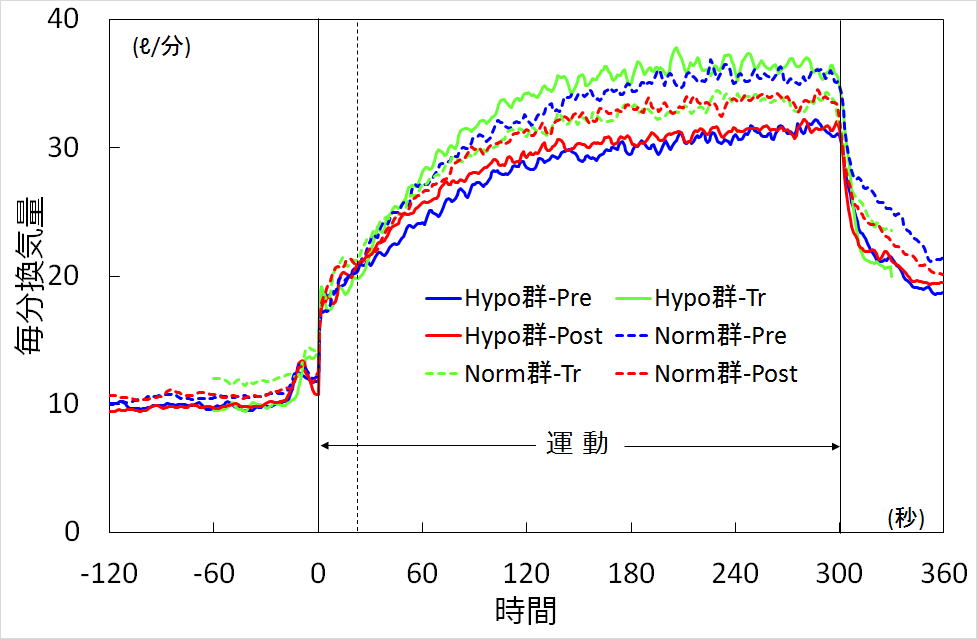
図2.低酸素トレーニング前後の毎分換気量の変化
時間相別に詳しく検討しますと,Norm群では,トレーニング期間中(Tr),Preに比べ毎分換気量の安静値からの変化分
(△VE)が,運動開始30秒から2分30秒までのPhase II及び3分50秒から4分50秒までの定常状態(Phase III)で10%程度
有意に低下しており,それは主に呼吸数の低下によって引き起こされていました.一方,Hypo群ではトレーニング期間中は
Preに比べPhase II, Phase IIIで△VEが20〜30%有意に増加しており,それは主に一回換気量の増大によって引き起こされて
いました.
PreとPostの比較では,Phase IIはNorm群で6%減少,Hypo群で6%増加し,Phase IIIではNorm群が8%減少に対しHypo群は
1%増加と,両群で反対の変化を示しますが,各群ともPre-Postで有意な変化は認められませんでした.
しかし,Phase II, IIIの△VEについて両群のPre-Postで交互作用が認められました.Phase Iで差は認められませんでしたが,
これはガス切替え直後は徐々に酸素濃度が低下するために,開始20秒では低酸素の影響がまだ少ないことが原因と推察されます.
結論
このように低酸素を用いて,被検者に気付かれずに運動時の換気を通常より増大させて短期間トレーニングさせると,
換気が過度に上がらない通常の条件(常酸素)に戻しても,換気を上げようとする「学習」が起こっていることが確認されました.
すなわち,運動時の換気亢進に「学習効果」が関与することが明らかとなりました.
(3)まとめ
2つの実験の結果は,運動時の呼吸が従来の神経性要因だけで決定されるのではなく,繰り返しの「学習」に
よっても影響を受けることを示唆しています.また,面白いことに,コントロール群ではPre に対しPostで換気量が低下し,
効率のよい呼吸が可能となっていることも一種の「学習」といえるかもしれません.
本研究は,運動時換気亢進の「学習仮説」を支持する有力な証拠となるとともに,運動時の呼吸が学習によって変容しうる
ことを明らかにした点で,非常に有意義であると言えます.
今後の展望として,換気を人為的に減少させた場合や,認知(学習)を妨げた場合についても明らかにする必要があります.
研究のページにもどる
トップページにもどる