細胞分子レベル研究
膵のアルカリ分泌の細胞・分子メカニズム
食事をすると、脳相、胃相、腸相をかいして、膵液の分泌が増えます。脳相は「パブロフの犬」によってよく知られていますが、最も大事なのは腸相で す。胃から十二指腸へ流れ込んだ胃酸、脂肪酸、アミノ酸が、十二指腸粘膜内にある内分泌細胞を刺激すると、cholecystokininや secretinといった消化管ホルモンが血中に放出されます。これらのホルモンが直接、あるいは自律神経系を介して膵臓の細胞に働くのです。
膵液は、高濃度(刺激時には140 mMに到達)の重炭酸イオン(HCO3-)を含むアルカリ性(pH8以上)の等張液(〜2 L/day)に消化酵素を混じたものです。アミラーゼやトリプシンなどの消化酵素は、膵臓の約90%をしめる腺房細胞から分泌されます。一方、HCO3-と水は、消化酵素を十二指腸へ運ぶ膵管系の上皮細胞(導管細胞)から分泌されます。
 (図1)
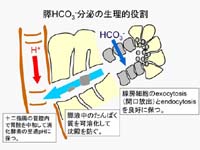
(図1)
このアルカリ分泌(HCO3-と水の分泌)によって、十二指腸内で胃酸が中和され消化酵素の至適pHに保たれることにより、栄養素の消化吸収が効率よく行われます。また、HCO3-と水の分泌は、膵臓内では、腺房細胞から分泌されるタンパク質を多く含んだ分泌液を、十二指腸内に向かって流し出しています。ですので、何らかの原因でHCO3-と水の分泌が不足すると、膵液が粘稠になり、タンパク栓や石ができて詰まることもあります。これが、嚢胞線維症や慢性膵炎の病態です。
私達の研究室では、この「膵導管細胞におけるHCO3-分泌メカニズム」を研究しています。実体顕微鏡で見ながら小動物(マウス、ラット、モルモット)の膵臓を針でほぐしていくと、
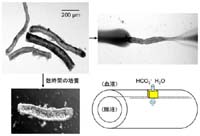 図2
図2
のような膵管(直径100ミクロン前後)を、“土管”の形を保ったまま単離することができます。膵管は膵臓全体の数%ぐらいですのでなかなか骨が折れる作
業ですが、慣れると10分間に1本ぐらいのペースで採れるようになります。膵管はほぼ均一な細胞(膵導管細胞)で構成された一層の上皮ですので、私達は、
これをバラバラに(single
cellに)することなく、そのまま実験に用いています。この方法の利点は、上皮膜としての構造(≒細胞の極性)と機能(この場合はHCO3-と水の分泌)が保たれた状態で、細胞レベルのシグナル(細胞内の各種イオン濃度、pH、電位)を測定することができることです。実際、単離した膵管を数時間培養すると、両端が自然に閉じて、閉鎖腔になった管腔がHCO3-と水の分泌によって膨らみます。また、管腔内を表層とは別個の組成の溶液で灌流することにより、生体内での状況をシュミレーションすることもできます。
両端が閉じた膵管を用いて、非常にシンプルでありながら重要な実験をすることができます。顕微鏡のステージ上で37℃の(精密な温度管理が必要です)HCO3--CO2緩衝液で表層灌流した時に、管腔が膨らむスピードを解析することによって、膵管上皮の単位表面積当たりの膵液分泌量を求めることができるのです(ビデオクリップを入れることができますか?)。
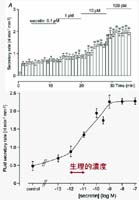 図3
図3
のようにsecretinの濃度を上げていくとともに溶液分泌量が増えます。食後の血中secretin濃度に相当する生理的濃度で十分な反応が見られるので、このpreparationの生理機能はほとんどintactだと言えます。
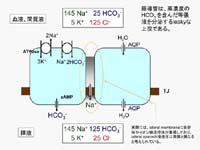 図4
図4
は、膵管上皮をかいするHCO3-と水の分泌の仕組みをごく簡略化して示したものです。血液から細胞内へのHCO3-の取り込みは、Na+の濃度勾配を利用してHCO3-を輸送するNa+-HCO3-
cotransportが担い、そのNa+濃度勾配はNa+-K+ ATPaseによって作られます。細胞内に蓄積されたHCO3-はcyclic
AMPによって活性化される陰イオン輸送体(CFTR Cl- channelを含む)を介して膵液中へ分泌されます。このHCO3-の能動輸送によって、経上皮的にわずかな浸透圧勾配が形成され、これを駆動力として、水が水チャネル(aquaporin)を通って血液中から膵液中へ移動します。また、電気的中性を保つために、輸送されたHCO3-と同量のNa+が
傍細胞経路(paracellular
pathway)を通って移動します。膵管上皮は、tight junctionの陽イオン透過性が高い(transepithelial
potentialが小さい、あるいはleakyな)上皮に分類されます。膵管上皮は、aquaporinが豊富に発現しているため水の透過性が高く、
paracellular
pathwayのNa+透過性も高いため、膵液は多量に分泌されながらも常に等張液になるのです。
膵導管細胞の特徴であり、私達の研究対象であるところは、非常に高濃度のHCO3-を分泌する点です。等張のNaHCO3水 溶液を分泌していると言ってもよいほどです。これは他の組織や細胞では見られません。最近の分子生物学の進歩によって、膵導管細胞に発現しているイオン輸 送体については、多くのことがわかってきました。しかし、どうも、膵導管細胞に特別なイオン輸送体が存在するわけではないようです。ですので、これらのイ オン輸送体が特殊な配置で局在するか、あるいは特殊な組み合わせで協調することによって、HCO3-分泌に特化していると推定されます。私達の研究室では、ヒトと同程度に高濃度のHCO3-を分泌するモルモットの膵臓から、上述のように小葉間膵管を単離して、(実験手法については、トップページの「実験方法」をクリックしてください。)、細胞内pH、Cl-濃度、電位を測定することによって、独自の細胞モデル
 (図5)
(図5)
を作成しました。ごらんになるとおわかりのように、このモデルは、生理学的ないくつかのイオン輸送機能から構築されており、それぞれの機能に相当する分子を特定していません。今後の課題は、この細胞モデルを、如何に分子の情報と結合させるか?です。
Created by MMS-NET